
Steve Vigdor, April 2, 2019
No scientific subject has garnered more persistent public resistance since its inception than evolution. Like big bang cosmology and the geologic time scale of the Earth, which we have dealt with here, evolution of the species cannot be reconciled with a literal reading of the origin story in the Judeo-Christian Bible, and so has been continually denied by creationists. But perhaps at an even more profound level, the concepts of evolution call into question the special nature of humans, and their relationship to a Creator, in the history of our planet. As a result, various religious groups and individuals have fought many court and political battles, continuing to this day, against the teaching of evolution in U.S. public schools. In recent decades those battles have centered on the demand that “creation science,” or its more modern church-detached descendant, Intelligent Design, be given equal time in school science curricula. In this blog series, we will deal with the extensive scientific evidence supporting current theories of evolution and the flaws in pseudoscientific counter-arguments offered by creationists and Intelligent Design advocates. The full blog series will follow the outline below:
Part I: Concepts and Controversy
1) A brief history
2) The tree of life
3) Controversy and court cases
4) The emergence of the Intelligent Design movement
5) Public opinion
Part II: Evolution in Action
6) Bacterial evolution in action
7) Antibody evolution
8) Fossil evidence
9) Biogeographical evidence
10) Objections to fossil evidence from Intelligent Design advocates
Part III: Genomic Evidence
11) DNA and DNA coding
12) DNA replication and the nature of mutations
13) Redundancy, robustness and natural selection forcing
14) Genomic similarities among different species
Part IV: Irreducible Complexity Reduced
15) Proposed examples of irreducible complexity
16) Flagellum regained
17) Evolution of the eye
18) Digital life confronts ID pseudoscience
19) Summary
We start with a brief history of the development of modern ideas of evolution.
1) A brief history
The idea that species evolve over time, and resistance to that idea, both began with the dawn of the 19th century. By the beginning of that century, the study of fossils had provided compelling evidence that a variety of animal species had grown extinct at some past time in Earth’s history. The fossil record, together with his study of living animal species, led French naturalist Jean Baptiste Lamarck to theorize that life on Earth was not fixed, but rather evolved naturally from simple toward increasingly complex forms as organisms adapted more successfully to their environment. This new concept of natural evolution was widely rejected at the time, especially by British naturalists who believed firmly that the variety of life on Earth reflected God’s benevolent design.
While some general concepts of Lamarck’s theory persist in the modern theory of evolution, two major detailed features of his work are considered incorrect, namely, that organisms evolve more or less linearly along a single ladder of complexity and that characteristics acquired by an individual during its lifetime could be passed along to its offspring. By the middle of the 19th century, Charles Darwin and Alfred Russel Wallace independently developed new concepts of evolution, according to which the development of new species was viewed as analogous to the branching of a tree, as opposed to Lamarck’s climbing of a ladder (see Fig. 1.1).

Based on their independent studies of many animal and plant species in a variety of geographical, often exotic, locales, Darwin and Wallace both conceived that living species had descended from common ancient progenitor species, but along tree-like branches that optimized the species’ adaptation to their particular environments. Both Darwin and Wallace were inspired by the late 18th century work of British cleric Thomas Malthus. In his book An Essay on the Principle of Population, Malthus had argued that unbridled human population growth would lead to an inevitable struggle over scarce resources because “the power of population is indefinitely greater than the power in the earth to produce subsistence for man.” Darwin and Wallace argued that plants and animals should be subject to similar struggles for resources, and that those with features best adapted to survive and breed would tend to pass those features along to offspring, and would come naturally to dominate the population. Darwin coined the phrase natural selection to describe this principle of evolution.
Darwin and Wallace both published some of their research in 1858, after decades of study, but it is the 1859 publication of Darwin’s ideas in book form that is viewed as the launch point for the field of evolutionary biology. The full title of the book (see Fig. 1.2) makes its central concept clear: On the Origin of Species by Means of Natural Selection, or Preservation of Favoured Races in the Struggle for Life. Darwin did not discuss the origins and evolution of mankind in this book, because he considered the subject too surrounded by prejudice and too devoid of fossil evidence. He waited until 1871, when he published The Descent of Man and Selection in Relation to Sex. In this book he argued that the available evidence was consistent with humans also having evolved from a common ancestor shared with apes and originating in Africa. He suggested that differences among human races might be influenced not only by natural selection, but also by sexual selection, i.e., by females exerting mating preferences for different traits among males of the species.

Meanwhile, Wallace established the scientific subfield of biogeography, typified by his 1876 book The Geographic Distribution of Animals. Here he argued that the different ranges found for different species reflected not only their adaptation by natural selection to different environments, but also their geographical isolation from other species with which they could, in principle, have interbred. The observed geographical distributions of animals have come to be more fully understood only in the latter half of the 20th century, in the framework of migration patterns influenced by historical continental drift associated with plate tectonics.
Darwin introduced the central concepts of common descent and natural selection into the language of evolution, but he did not gain an understanding of the mechanism for inheritance of naturally selected traits. The first hints of genetic inheritance, and of the existence of dominant and recessive variants of genes, came from experiments carried out by Gregor Mendel around the same time that Darwin was publishing Origin of Species. Mendel experimented with hybrids of different species of peas, some characterized by smooth skin and others by wrinkled texture. While the first generation hybrids all exhibited smooth texture, one-quarter of the second generation peas produced from those hybrids again had wrinkled texture. But the significance of Mendel’s result was not understood until the discovery at the end of the century – fifteen years after Mendel’s death – of chromosomes within the nucleus of each animal or plant cell.
The chromosomes contain sections labeled genes that determine an individual’s traits. It was found that when sperm fertilize eggs to create offspring, each parent contributes half of the full complement of chromosomes. Human cells, for example, contain 23 pairs of chromosomes, with each parent contributing one chromosome to each pair. The human chromosomes comprise a total of more than 20,000 genes. Each individual born via sexual reproduction contains two copies, although often different variants, of each gene. Some variants of particular genes lead to dominant traits, while other variants are recessive and remain masked in the traits of the individual. But when two individuals mate, if each carries a dominant variant with a recessive variant of the same gene, there is a 25% probability that the offspring will get the recessive variant from each parent, so that the formerly masked trait reappears in the next generation. This realization explained the findings of Mendel and of other researchers who found similar 3-to-1 ratios among offspring in experiments near the end of the 19th century. The variants of a particular gene are known as alleles.
Scientists speculated that the mechanism for Darwin’s evolution of adaptation might be associated with occasional errors in gene duplication during reproduction, so-called mutations. The occurrence of such genetic mutations was demonstrated in experiments on fruit fly evolution, carried out in the Columbia University laboratory of Thomas Hunt Morgan. Every fly in one particular evolutionary line was born with red eyes, until a single individual emerged with white eyes, signaling a sudden genetic mutation. When Morgan mated the white-eyed fly with a red-eyed fly, he produced a generation of red-eyed hybrids, since the red pigmentation resulted from a dominant allele. But when he mated members of that first new generation, the by then familiar one-quarter of the grandchildren had white eyes, signaling that the mutated allele was being inherited. The work of Morgan and contemporaries launched the new science of genetics, and Morgan himself was awarded the 1933 Nobel Prize in Physiology or Medicine “for his discoveries concerning the role played by the chromosome in heredity.”
Morgan had demonstrated the hereditary persistence of a genetic mutation, but had not created a new species. The white-eyed fly was still fully capable of mating with the red-eyed flies. Subsequent geneticists demonstrated that any given trait was generally determined by many genes, and that a mutation to a single gene could create small changes, rather than drastic transformations, of traits. But they also began to build sophisticated mathematical models of evolution in a large population, which made it plausible that natural selection could indeed gradually propagate a mutation, or series of mutations, that offered even a slight advantage in fitness throughout a population over many generations. The Soviet-born geneticist Theodosius Dobzhansky then demonstrated that isolated populations of fruit flies, allowed to propagate different mutations across many generations, eventually produced populations that were unable to cross-breed, or if they did were unsuccessful in producing hybrids. Dobzhansky realized that this evolved inability to cross-breed was the crucial criterion in producing new species. He described this process of speciation, illustrated in Fig. 1.3, in his 1937 book Genetics and the Origin of Species, and this work helped to establish the so-called Modern Synthesis of evolutionary theory.

An understanding of the mechanisms for genetic mutation at the biomolecular level began in earnest only after the 1953 discovery by Francis Crick and James Watson, based upon X-ray diffraction images that had been obtained by Rosalind Franklin, of the double helix structure of DNA, the complex biomolecule in chromosomes. Subsequent work revealed how DNA molecules encode the instructions for synthesizing proteins and other complex biomolecules essential for a variety of biological functions. It also revealed that many possible mutations that can occur during DNA replication in cell division are neutral, with no effect on biological function. Andreas Wagner and collaborators at the University of Zurich have shown that the plethora of such neutral mutations generates an enormously expanded network of possible multi-step pathways that may lead eventually to additional advantageous mutations to be favored by natural selection. Wagner has described how this multiplicity of pathways aids evolution in his 2014 book Arrival of the Fittest: Solving Evolution’s Greatest Puzzle (you can also see a video of Wagner presenting these ideas here). And the 21st century explosion in the technology for mapping the DNA details in a variety of species has provided an enormous quantity of biomolecular evidence in support of the modern theory of evolution, some of which we will discuss in later sections.
It has been clear throughout the history of genetics that mutations and natural selection do not provide the only mechanism for generating genetic differences among populations. A competing mechanism is genetic drift. This is a process whereby the random selection of some alleles over others during reproduction may enhance the probability of finding those alleles in subsequent generations, and lead eventually to their complete dominance, even though there may be no adaptive advantage gained by that dominance.

Figure 1.4 illustrates how genetic drift can occur randomly for a small population with initially equal frequencies of occurrence for two distinct alleles (represented by red and blue marbles in the figure) of a single gene. The transition from one jar to the next in the figure is intended as a schematic for a new generation, formed when 20 organisms from the current jar are selected at random to reproduce asexually, yielding one clone in the next jar. In this random selection, it is entirely possible for a single organism to be selected multiple times, so that it produces multiple clones in the next generation. Although there are equal probabilities to choose a red or a blue marble from the first generation, there are many more unbalanced combinations in the choice of 20 than the single balanced combination of 10 blue marbles and 10 red marbles. Hence, it is likely that the second generation will, purely by chance, contain different frequencies of occurrence for the red and blue alleles. The figure illustrates one particular possibility, with 14 blue and 6 red marbles in the second jar.
The randomly generated dominance of blue alleles in the second generation in Fig. 1.4 increases the probability for selecting blue marbles for cloning to produce the third generation, and so forth. After some number of generations, this random process will lead to complete dominance of one allele (blue in the figure, but could equally well have been red in an independent trial) and the disappearance of its alternative. Although the illustration was made for cloning reproduction, the probabilistic mechanism for genetic drift applies equally well to sexual reproduction processes, in which each parent contributes an allele.
It was understood throughout the 20th century that genetic drift could play a significant role within small, isolated populations. It is the mechanism at work, for example, in the high rate of occurrence of some rare conditions among inbred populations, such as Ellis-van Creveld syndrome among Amish communities in America or total rod cell color blindness on the Pingelap atoll in Micronesia. Over the past half-century, however, it has become clearer that neutral mutations and genetic drift often combine to generate genetic differences among even large, but geographically separated, populations within a given species.
These, then, are the central concepts of modern evolutionary theory: common descent, natural selection, genetic mutations, genetic drift. Darwin’s concept of sexual selection, introduced in The Descent of Man, is viewed as one aspect of natural selection, an aspect that sometimes favors attraction of a mate for reproduction over even protection against predators or survival of the individual. Sexual selection has been emphasized recently in considerations of the evolution of beauty among living organisms.
2) The Tree of Life
A century-and-a-half of detailed research on anatomical and genomic (i.e., mapped DNA) similarities and differences among distinct living species, together with extensive fossil evidence for extinct species, has fleshed out Darwin’s concept of evolution along the branches of a tree, spreading from a universal common ancestor. The consensus tree of life, or phylogeny, of the species is illustrated in its broad outlines in Fig. 2.1, where successive indicated branch points are understood to have occurred chronologically during Earth’s lifetime.

Any one of the broad branches included in Fig. 2.1 can be magnified to illustrate the separations among more closely related species. For example, Fig. 2.2 illustrates our best current understanding of the evolutionary tree of the primates, this time with the timeline explicit. The closest relatives to humans, revealed in great detail from chromosomal similarities, are the chimpanzees. The separate developments of chimps and humans are believed to have branched off from a common ancestor some 6—8 million years ago. While we do not yet have fossils of the purported common ancestor, we have fairly detailed fossil evidence of the subsequent stages of evolutionary development of humans (genus homo), as will be discussed in Part II of this blog series.

The modern era of mapping complete genomes for different species, in full biomolecular complexity, has led to remarkable resolution in phylogenetics, as illustrated by the fantastically detailed tree of life in Fig. 2.3.

The theory of evolution does not encompass the question of how the common ancestor or ancestors – the first life forms on Earth, preceding branch point C in Fig. 2.1 – came to be. Darwin himself speculated on this in an 1871 letter to Joseph Hooker: “But if (and oh what a big if) we could conceive in some warm little pond with all sorts of ammonia and phosphoric salts, light, heat, electricity, et cetera present, that a protein compound was chemically formed, ready to undergo still more complex changes…” The currently preferred speculations about the origin of life, referred to as abiogenesis, are not so far from Darwin’s: they propose that the first complex biomolecules – perhaps simple forms of RNA – capable of storing genetic information and undergoing self-replication were assembled by chance, over hundreds of millions of years in water that also contained mineral surfaces, from simple non-living organic compounds formed in earlier eons. But this question remains a subject so far of speculations, with supporting experimental evidence hard to come by. However, the theory of evolution does not require an answer to this origin question; evolution can be judged on its own merits, with many levels of supporting observational evidence.
3) Controversy and Court Cases
Fundamentalist religions that regard the Bible as a literal record of Earth’s history have long viewed evolutionary science as an existential threat. The concepts of universal common descent and evolution over billions of years via genetic mutations and natural selection stand in stark contrast to the biblical creation of all living species by God on days 5 and 6 of “creation week.” As we have detailed in our blog series on Young Earth Creationism, creationists accept that there is a fossil record of species extinctions, but they attribute those extinctions to the Great Flood. They view the deposition of distinct fossils in different geological strata not as reflections of a chronological record of evolution, but rather as an indication that some co-existing species were better able than others to outrun the rising flood waters in the race to higher ground. They view the concept that humans and apes descended from common ancestors as repulsive. Creationists view science in conflict with the biblical account of creation as direct threats to the moral authority of their religion.
In the 1920s in the U.S., as the modern synthesis of evolutionary theory was taking root, there was an upsurge of fundamentalist religious fervor. This led to a movement to prevent the teaching of evolution in public schools and to remove it from biology textbooks. Several state legislatures, including South Carolina, Oklahoma, Kentucky and Tennessee passed laws prohibiting the teaching of evolution. The Tennessee law was named for State Representative John Butler, who was the head of the World Christian Fundamentals Association. Butler later admitted, “I didn’t know anything about evolution…I’d read in the paper that boys and girls were coming home from school and telling their fathers and mothers that the Bible was all nonsense.” The Butler Act was the first of the anti-evolutionary laws to be tested in court, in the famous 1925 Scopes “Monkey” Trial, later immortalized in the play and movie Inherit the Wind.
The Scopes trial was a spectacle, financed by the American Civil Liberties Union in order to test the constitutionality of the law and held in the small town of Dayton, Tennessee, with nationwide radio coverage. It pitted populist Fundamentalist William Jennings Bryan, a three-time presidential candidate, for the prosecution, arguing against famed defense lawyer and agnostic Clarence Darrow. The defendant was John T. Scopes, a Dayton high school science and math teacher who wasn’t sure he had, in fact, ever taught evolution, but nonetheless agreed on principle to serve as defendant and coached students to testify against him. The presiding judge, John T. Raulston, opened the trial quoting the Book of Genesis and the Butler Act, warned the jury not to judge the merit of the law, but only the violation of the Act, and disallowed most of the witnesses the defense wanted to call to testify, judging them to be irrelevant to the case. Bryan spoke against evolution for teaching children that humans were but one of 35,000 types of mammals, and specifically argued that the theory claimed humans were descended “not even from American monkeys, but from old world monkeys.” One of the defense lawyers, Dudley Malone, warned of inquisitions and argued that the Bible should be preserved in the realm of theology and morality and not put into a course of science.
The culmination of the trial was defense lawyer Darrow calling prosecution attorney Bryan to testify as an expert on the Bible. The session was held outside on the courthouse lawn, with the jury sequestered. Darrow asked Bryan about various supernatural occurrences reported in the Bible, and told him at one point, “You insult every man of science and learning in the world because he does not believe in your fool religion.” Bryan, in his turn, insisted that the Bible be quoted verbatim rather than allowing Darrow to paraphrase it for his own purposes. After two hours of examination, the judge abruptly adjourned the court, ruled that the entire examination was irrelevant to the case and should be expunged from the court record, and denied Bryan a chance to cross-examine the defense lawyers in return. The jury took only nine minutes of deliberation to find Scopes guilty and the judge ordered him to pay a $100 fine.
No closing arguments were delivered to the jury in the Scopes trial by either the defense or the prosecution. But Bryan distributed his intended summation to reporters, and it provides a flavor of the creationist argument at that period, having essentially nothing to do with the science of evolution or the factual evidence supporting it:
“Science is a magnificent force, but it is not a teacher of morals. It can perfect machinery, but it adds no moral restraints to protect society from the misuse of the machine. It can also build gigantic intellectual ships, but it constructs no moral rudders for the control of storm-tossed human vessel. It not only fails to supply the spiritual element needed but some of its unproven hypotheses rob the ship of its compass and thus endanger its cargo. In war, science has proven itself an evil genius; it has made war more terrible than it ever was before. Man used to be content to slaughter his fellowmen on a single plane, the earth’s surface. Science has taught him to go down into the water and shoot up from below and to go up into the clouds and shoot down from above, thus making the battlefield three times as bloody as it was before; but science does not teach brotherly love. Science has made war so hellish that civilization was about to commit suicide; and now we are told that newly discovered instruments of destruction will make the cruelties of the late war seem trivial in comparison with the cruelties of wars that may come in the future. If civilization is to be saved from the wreckage threatened by intelligence not consecrated by love, it must be saved by the moral code of the meek and lowly Nazarene. His teachings, and His teachings alone, can solve the problems that vex the heart and perplex the world.
It is for the jury to determine whether this attack upon the Christian religion shall be permitted in the public schools of Tennessee by teachers employed by the state and paid out of the public treasury. This case is no longer local, the defendant ceases to play an important part. The case has assumed the proportions of a battle-royal between unbelief that attempts to speak through so-called science and the defenders of the Christian faith, speaking through the legislators of Tennessee. It is again a choice between God and Baal; it is also a renewal of the issue in Pilate’s court.”
William Jennings Bryan died five days after the end of the trial. The defense lawyers appealed Scopes’ conviction to the Supreme Court of Tennessee, which rejected their arguments based on ways in which the Butler Act violated aspects of the U.S. and Tennessee State Constitutions. However, the court set aside the conviction on a technicality, because the judge had ordered a $100 fine, while the State Constitution insisted that only juries could set fines above $50. The court decision insisted that nothing was “to be gained by prolonging the life of this bizarre case.” Consequently, there was no retrial, no appeal to higher courts, and no definitive decision about the constitutionality of anti-evolutionary laws.
In the wake of the Scopes trial, creationists succeeded in getting 13 other states, both northern and southern, to consider anti-evolution laws, but they passed state legislatures only in Mississippi and Arkansas. Nonetheless, their efforts had the desired effect of suppressing the teaching of evolution in many U.S. schools. The Arkansas law was eventually declared unconstitutional by the U.S. Supreme Court in the 1968 case Epperson v. Arkansas. The Supreme Court held that the Arkansas statute violated the Establishment Clause of the First Amendment, which prohibited states from requiring “that teaching and learning must be tailored to the principles or prohibitions of any religious sect or dogma.”
In the wake of the Soviet launch of Sputnik, and thereby of the space race, in 1957, science education in the U.S. took on greater urgency and prestige. The teaching of evolutionary theory and its supporting evidence was reintroduced as a standard aspect of science curricula. This led creationists to shift their tactics in the 1960s from an anti-science to a pseudoscience stance. They began to focus on what they called creation science, based on less overtly religious arguments and employing questionable claims of scientific evidence said to support the literal interpretation of the Bible. As we have detailed elsewhere on this site, the claims of creation science in support of a young Earth, and in stark opposition to an overwhelming base of scientific evidence, are misleading, incoherent, supernatural and unconvincing. They represent pseudoscience invented after the fact to justify what creationists viewed as an infallible account dictated by the Bible. Hoping to attract to themselves some of the new-found prestige for science, they tried to demand that their creation science should be taught alongside the theory of evolution in public schools.
In the early 1980s several states, including Arkansas and Louisiana, used a template prepared by the Institute for Creation Research to pass “balanced treatment” laws requiring that if evolutionary science was taught in public schools, creation science had to be taught as well, as an alternative scientific theory. In the 1982 case McLean v. Arkansas Board of Education, a U.S. District Court ruled the Arkansas balanced treatment statute to be unconstitutional as it again violated the Establishment Clause of the First Amendment of the U.S. Constitution. In his ruling, Judge William Overton classified creation science as religion, since it failed to meet what he set out as the essential characteristics of any science: (1) it is guided by natural law; (2) it has to be explanatory by reference to natural law; (3) it is testable against the empirical world; (4) its conclusions are tentative, i.e., are not necessarily the final word; and (5) it is falsifiable. Overton summarized the case against creationism as science in the following quote from the ruling: “While anybody is free to approach a scientific inquiry in any fashion they choose, they cannot properly describe the methodology as scientific, if they start with the conclusion and refuse to change it regardless of the evidence developed during the course of the investigation.”
Overton’s ruling was binding only on schools within the Eastern District of Arkansas. A national standard was subsequently set in the 1987 U.S. Supreme Court case Edwards v. Aguillard judging the balanced treatment statute passed in Louisiana. In a 7-to-2 decision (with Justices Antonin Scalia and William Rehnquist dissenting), the Court found that the Louisiana act represented an unconstitutional infringement on the Establishment Clause because it failed a three-prong test that required legitimate government actions to (1) have a legitimate secular purpose, (2) not have the primary effect of either advancing or inhibiting religion, and (3) not result in an “excessive entanglement” of the government and religion. However, the ruling written by Justice William Brennan did leave a loophole through which creationists would next try to wriggle: “We do not imply that a legislature could never require that scientific critiques of prevailing scientific theories be taught. . . . Teaching a variety of scientific theories about the origins of humankind to schoolchildren might be validly done with the clear secular intent of enhancing the effectiveness of science instruction.”
4) The Emergence of the Intelligent Design Movement
Prior to the Edwards v. Aguillard case reaching the Supreme Court, creationists had begun work on a creationist biology textbook, with the initial title Creation Biology. The book advanced through several drafts and titles as the Supreme Court case approached, and then in the wake of the Court decision. It was finally published in 1989 by the Texas-based Christian non-profit organization Foundation for Thought and Ethics, with the title Of Pandas and People. As revealed in Fig. 4.1 the drafts went through a phase transition in the wake of the Supreme Court 1987 ruling, replacing the word “creation” throughout with “intelligent design” (a phrase the book’s editor Charles Thaxton “picked up from a NASA scientist”) and “creationist” with “design proponent.”

For example, the 1986 draft contained the following definition: “Creation means that the various forms of life began abruptly through the agency of an intelligent creator with their distinctive features already intact. Fish with fins and scales, birds with feathers, beaks, and wings, etc.” In the later drafts this was slightly modified to read: “Intelligent design means that various forms of life began abruptly through an intelligent agency, with their distinctive features already intact. Fish with fins and scales, birds with feathers, beaks, wings, etc.” Thus, the Intelligent Design (ID) movement was born.
The published version of Of Pandas and People backs off from pushing the debunked arguments for Young Earth Creationism, commenting that some “take the view that the earth’s history can be compressed into a framework of thousands of years, while others adhere to the standard old earth chronology.” But the book raises numerous objections to the theory of evolution – objections we will debunk in later sections of this blog series – and concludes that anatomical features of living species are too complex to have evolved gradually. Rather, it claims that these features must have been the product of intelligent design, although the book refrains from explicitly connecting the designer to the Christian God, thereby hoping to avoid dismissal via the Establishment Clause of the Constitution. The book also fails to explain why “life beginning abruptly” should be viewed as an inherently more “intelligent” design than giving organisms the capability to adapt gradually through genetic change to a delicately balanced environment that is itself subject to continual change.
The publishers worked assiduously throughout the 1990s to get Of Pandas and People adopted as a biology textbook in U.S. schools. They mobilized local Christian conservative groups to push school boards and teachers on the subject, and to run for school board and local educational committee positions themselves. Their efforts were aided by the 1990 establishment of the Discovery Institute by Bruce Chapman, an organization that has become the most prominent proponent of ID ideas. The Discovery Institute began a campaign to “teach the controversy” by presenting concepts of evolution and intelligent design side by side in public schools. They all met with little success, despite even a 2001 (failed) attempt by Senator Rick Santorum of Pennsylvania to add a “Teach the Controversy” amendment to that year’s federal Education funding bill.
But finally, in October 2004 the Dover Area School District School Board in York County, Pennsylvania voted 6-3 to amend the district’s science curriculum to include Intelligent Design, after Board member William Buckingham announced that he’d received a donation of 50 copies of the second edition of Of Pandas and People to use as reference books. This decision was met very shortly thereafter by a lawsuit filed by the ACLU, Americans United and Pepper Hamilton LLP on behalf of Tammy Kitzmiller, the mother of a ninth grade Dover biology student. Judge John E. Jones III presided over the 2005 Kitzmiller v. Dover Area School District case. Jones issued a 139-page “findings of fact” and ruled that “ID is nothing less than the progeny of creationism” and is not science. He permanently barred the Dover School Board from “maintaining the ID policy in any school within the Dover Area School District, from requiring teachers to denigrate or disparage the scientific theory of evolution, and from requiring teachers to refer to a religious, alternative theory known as ID.” In November 2005, the 8 members of the Dover School Board who were up for re-election all lost to challengers who opposed teaching ID in science classes. The new School Board refused to appeal the Kitzmiller ruling.
Stephen Meyer, one of the most prominent advocates for ID, insists that Intelligent Design is not Creationism, but Fig. 4.1 and the courts beg to disagree. Yet despite their inability to convince any courts or many scientists or school boards, ID limps onward, continuing to push school districts to “teach the controversy.” The movement systematically misrepresents the extensive evidence (to be discussed in Parts II and III of this blog series) supporting the modern synthesis of evolutionary theory, and it rests on religious faith, one self-serving philosophical argument and one more recently enunciated pseudoscientific principle.
The philosophical argument was originally framed by Philip E. Johnson, a retired University of California law professor, one of the founders of the ID movement and a co-founder of the Discovery Institute’s Center for Science and Culture. Johnson terms his philosophy theistic realism. It maintains that “science, by relying on methodological naturalism, demands an a priori adoption of a naturalistic philosophy that wrongly dismisses out of hand any explanation that contains a supernatural cause.” Well, yes. Supernatural causes, by definition, cannot be questioned, explained or subjected to falsifiability tests. They fall outside the scientific method. If they were to be included, science would no longer be distinguishable from religion. So why not simply admit that ID is a religious concept, rather than trying so hard to pass it off as science?
The pseudoscientific principle at the heart of ID is irreducible complexity, a term introduced by Michael Behe in a chapter of the second edition of Of Pandas and People and central to Behe’s 1996 book Darwin’s Black Box: The Biochemical Challenge to Evolution (see Fig. 4.2). Behe is a Professor of biochemistry at Lehigh University and a Senior Fellow at the Discovery Institute Center for Science and Culture. Behe defines an irreducibly complex system as one “which is composed of several well-matched, interacting parts that contribute to the basic function, and where the removal of any one of the parts causes the system to effectively cease functioning.” He uses a mousetrap as an example. Behe claims that many such biological systems could not possibly result from multi-step genetic mutations, but can only have been produced whole by an intelligent design agent. As we will argue in Part IV of this blog series, the logic behind this claim is flawed and several of the specific examples Behe offers have been disproven. In particular, we will discuss a falsifiability test Behe proposed for the concept of irreducible complexity, a test that has indeed been carried out and shown the concept to be false. The concept was rejected by the National Academy of Sciences in the Kitzmiller case. That has not, of course, persuaded ID proponents to stop claiming irreducible complexity as the strongest evidence in favor of intelligent design.

A variant of irreducible complexity proposed by William Dembski is specified complexity. Dembski argued that another clear signature of intelligent design is the occurrence of biological patterns that are simultaneously specified – that is, easily described – and complex, that is, unlikely to occur by chance. The one example he offers is one also highlighted by Behe, the bacterial flagellum of E. coli. He specifies the description of this structure as a “bidirectional rotary motor-driven propeller,” and speculates that the probability of achieving such a four-basic-concept complex structure by a sequence of elementary logic operations is smaller than one part in 10140. Specified complexity is, in fact, a poorly formulated attempt to make a probabilistic argument against evolution by successive mutations, an attempt that has been roundly criticized as both muddled and grossly inaccurate by numerous experts in probability and information theory. Dembski has responded to critics by claiming that he is not “in the business of offering a strict mathematical proof for the inability of material mechanisms to generate specified complexity.”
5) Public Opinion
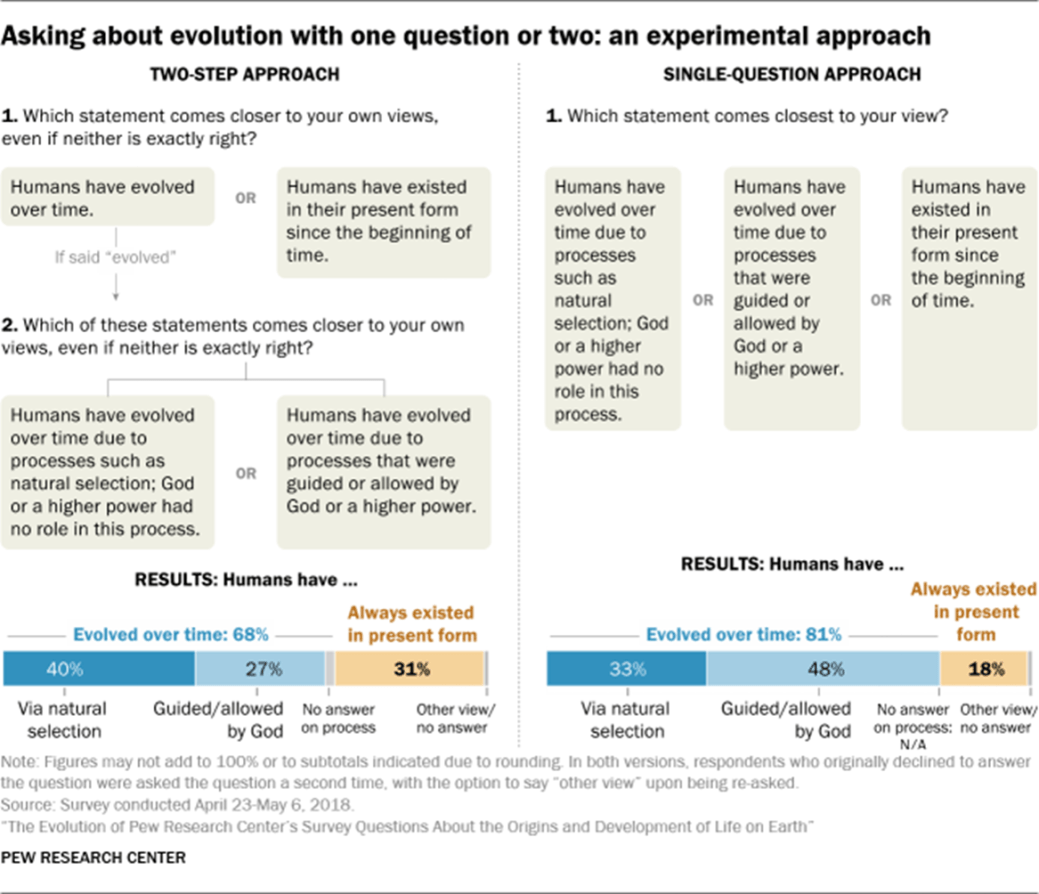
By now, evolution via natural selection has been taught nearly universally in U.S. public schools for decades, and ID advocates have met with very little success in getting school districts to “teach the controversy.” It is then interesting to investigate U.S. public opinion on the subject. A great deal of polling has been done on attitudes toward evolution. As illustrated in Fig. 5.1, the Pew Research Center has determined that the responses one gets depend on how the questions are posed. But independent of whether people are asked about human development via two successive questions with two choices apiece, or via a single question with three options, no more than 40% of all American respondents see evolution by natural selection as the primary mechanism. The good news is that more than two thirds of respondents appear to accept the idea of evolution, although many believe that evolution is guided by God rather than by natural selection of fitness for survival and reproduction. Despite extensive fossil evidence to the contrary (to be discussed in Part II), somewhere between one-fifth and one-third of respondents believe that humans have existed in their present form since the beginning of time.
Not surprisingly, the responses are strongly correlated with religious affiliation. The results in Fig. 5.2 come from a 2014 Pew survey using the two-question approach on the left side of Fig. 5.1. They indicate that well over half of evangelical Christians, of Mormons and of Jehovah’s Witnesses adhere to the belief that humans have always existed in their present form. 11% or less of these particular groups put credence in evolution by natural processes. Other religious groups and those unaffiliated with any organized religion are far more willing to accept the reality of evolution. Roughly one-third of Catholics, for example, believe in evolution guided by a supreme being, a stance well aligned with the official guidance of the Catholic Church, which accepts that the Bible cannot be taken as a literal record of history or science.

Finally, it is worth noting that religious affiliation seems to introduce a significant correlation between attitudes toward evolution and toward environmental regulation, reflective of a broader correlation between religious and political identification in the U.S. The correlation is illustrated in Fig. 5.3, assembled by Josh Rosenau from a massive Pew Religious Landscape Survey carried out in 2007. Rosenau examined the responses to two specific binary questions from that survey. The first asked respondents which of the following two statements they most agreed with:
a) Stricter environmental laws and regulations cost too many jobs and hurt the economy;
b) Stricter environmental laws and regulations are worth the cost.
The second question asked respondents whether they agreed or disagreed with the following statement:
Evolution is the best explanation for the origins of human life on earth.

Rosenau plotted the mean result for various religious groups to each question on a scale from -1 to +1, and represented the results by circles of areas proportional to the percentage of the 36,000 respondents belonging to each group. Note the strong correlation in Fig. 5.3, with the exception of the outlier responses from the small group of Jehovah’s Witnesses. In particular, the various evangelical Christian groups mostly occupy the lower left corner of the graph, representing strong opposition to both evolution and environmental regulation. In particular, we have shown polling elsewhere on this site that emphasizes the strong correlation between evangelical fundamentalism and denial of the reality of climate change and the role of humans in causing it. A willful dismissal of scientific evidence, even if motivated by otherwise laudable religious faith, can be quite dangerous to public health and welfare.
In the remaining parts of this blog series, we will discuss some of the extensive observational evidence now available supporting genetic mutation, evolution by natural selection, and universal common descent. We will furthermore discuss the serious flaws in creationist or intelligent design arguments opposing the modern synthesis of evolutionary theory.
— To be continued in Part II —
References:
https://evolution.berkeley.edu/evolibrary/article/0_0_0/history_01
https://science.howstuffworks.com/life/evolution/evolution.htm
https://en.wikipedia.org/wiki/Natural_selection
https://en.wikipedia.org/wiki/On_the_Origin_of_Species
https://en.wikipedia.org/wiki/The_Descent_of_Man,_and_Selection_in_Relation_to_Sex
https://en.wikipedia.org/wiki/Biogeography
https://en.wikipedia.org/wiki/Genetic_drift
https://en.wikipedia.org/wiki/Chromosome
https://en.wikipedia.org/wiki/Speciation
https://en.wikipedia.org/wiki/Genetics_and_the_Origin_of_Species
A. Wagner, Arrival of the Fittest: Solving Evolution’s Greatest Puzzle (Current, 2014) (https://www.amazon.com/Arrival-Fittest-Solving-Evolutions-Greatest/dp/1780745168/ref=cm_cr_arp_d_product_top?ie=UTF8)
http://www.talkorigins.org/faqs/comdesc/
I. Letunic and P. Bork, Interactive Tree of Life (iTOL): an online tool for phylogenetic tree display and annotation, Bioinformatics Advance Access (2006)(https://itol.embl.de/help/17050570.pdf)
https://en.wikipedia.org/wiki/Scopes_Trial
W.J. Bryan’s Summation of the Scopes Trial (https://www.beliefnet.com/News/1999/12/Faith-Of-Our-Fathers.aspx)
https://en.wikipedia.org/wiki/Epperson_v._Arkansas
https://en.wikipedia.org/wiki/McLean_v._Arkansas
https://en.wikipedia.org/wiki/Intelligent_design
https://en.wikipedia.org/wiki/Edwards_v._Aguillard
https://supreme.justia.com/cases/federal/us/482/578/
https://en.wikipedia.org/wiki/Of_Pandas_and_People
https://en.wikipedia.org/wiki/Teach_the_Controversy
https://en.wikipedia.org/wiki/Kitzmiller_v._Dover_Area_School_District
Stephen Meyer, Intelligent Design is Not Creationism (https://www.discovery.org/a/3191/)
Michael Behe, Darwin’s Black Box (Free Press, 1996) (https://en.wikipedia.org/wiki/Darwin%27s_Black_Box)
https://en.wikipedia.org/wiki/Theistic_science
https://en.wikipedia.org/wiki/Specified_complexity
http://www.pewresearch.org/science/
http://www.pewresearch.org/fact-tank/2017/02/10/darwin-day/
www.pewforum.org/essay/darwin-in-america/
J. Rosenau, https://ncse.com/blog/2015/05/evolution-environment-religion-0016359
